SYNTHESE
CHLOROPHYLLIENNE
Rudiments
Gérard
Gomez
Plan de l'étude
2) La phase claire de la photosynthèse
2-1)
Photosystème I et photosystème II
2-2)
Schéma synoptique des opérations de la phase claire
3) La synthèse des glucides ; Le cycle de Calvin
Annexe 1 NADP+/NADPH - ATP/ADP
Annexe 2 Chloroplastes, thylakoïdes, stroma
Annexe 3 Caroténoïdes – Carotènes- xanthophylles
Annexe 4 Chlorophylle A, chlorophylle B, xantophylles
Annexe 5 Valeurs des potentiels rédox des couples
intervenant dans la photosynthèse
|
Les
plantes vertes tirent l’énergie dont elles ont besoin pour vivre et se
développer de la lumière du soleil et produisent des substances, à partir du
dioxyde de carbone puisé dans l’air et à partir de l’eau, selon l’équation
suivante : |
|

Parce
que la formule brute de la plupart de ces substances, fait apparaître hydrogène
et oxygène dans les mêmes proportions que dans l’eau K.Schmidt leur a donné en 1844 le nom d'hydrates
de carbone. A cause de la saveur particulière de quelques
unes on les a aussi appelées sucres ; on continue parfois à employer ce
terme, mais on utilise plutôt aujourd'hui l'appellation glucides.
On
peut distinguer deux phases dans la photosynthèse :
- La phase claire
ou phase photochimique
C'est
l'action de la lumière ; elle a lieu dans les thylakoïdes des chloroplastes
(voir annexe 2). Les photons absorbés par la
chlorophylle permettent l'oxydation de l'eau ainsi que la libération d'ATP et
de NADPH qui vont donner respectivement l'énergie et un certain potentiel
réducteur, nécessaires à la synthèse du glucose et autres molécules organiques
lors de la phase obscure,.
- La phase obscure
ou phase biochimique
Elle
a lieu dans le stroma (fluide qui entoure les thylakoïdes) des chloroplastes
chez les eucaryotes. Il s'agit de réactions biochimiques conduisant du CO2
aux molécules organiques (glucose, fructose, saccharose …)
Ce
sont surtout les feuilles qui contiennent les chloroplastes et donc qui sont le
siège de la photosynthèse, mais on peut en trouver aussi dans les tiges de
certains végétaux.
Les
glucides ainsi synthétisés sont ensuite répartis dans toute la plante.
2) La phase claire
de la photosynthèse
La
phase claire correspond à des réactions photochimiques, permettant au travers
de deux photosystèmes PS I et PS II, le transport d'électrons pour aboutir à la
synthèse d'ATP et de NADPH.
2-1) Photosystème
I et photosystème II
La
phase claire a lieu grâce aux pigments contenus dans les membranes des
thylakoïdes : chlorophylle A, chlorophylle B, xanthophylles notamment (voir annexe 4).
La
chlorophylle A est le plus important de ces pigments ; unie à différentes
protéines elle constitue deux complexes moléculaires P 680 et P 700 dont l'absorption
optique a lieu respectivement à 680 nm et 700 nm. Ces zones d'absorption sont
différentes des pics d'absorption de la chlorophylle A seule.
Les
autres pigments (on dit qu'ils sont accessoires) absorbent la lumière à de plus
faibles longueurs d'onde (photons d'énergie supérieure). Ils transmettent une
partie de cette énergie aux molécules de chlorophylle A. De cette façon les
pigments accessoires renforcent l'action de celle-ci.
On
est en présence de deux antennes collectrices de photons.
L'énergie
des photons est ensuite transmise de molécule en molécule, par résonance,
jusqu'à un centre réactionnel qui contient une chlorophylle A particulière qui
une fois excitée a la propriété de céder un électron à un accepteur qui est de
ce fait réduit.
C'est
là que l'énergie lumineuse est transformée en énergie chimique. L'électron se
transmet de molécule en molécule au travers de molécules acceptrices
d'électrons, jusqu'à un état stable.
Il
existe deux centres réactionnels avec antennes collectrices d'énergie lumineuse
appelés photosystèmes I et II (PS I et PS II).
Remarque :
La
molécule de chlorophylle A excitée qui cède un électron à un accepteur, se
charge positivement. Grâce à un donneur d'électrons elle récupère rapidement
l'électron perdu et se régénère prête à recevoir de nouveau de l'énergie
lumineuse de la part de l'antenne collectrice.
2-2) Schéma synoptique des opérations de la phase claire
Le PS II a comme donneur
d'électrons, le complexe COE (Complexe d'oxydation de l'eau) et comme premier
accepteur d'électrons la phéophytine a.
Le schéma simplifié ci-dessous correspond
à un transfert acyclique d'électrons qui va de l'eau (donneur primaire
d'électrons) au NADP+ (accepteur d'électrons et de protons) au
travers des deux systèmes PS I et PS II.
C'est un schéma dit en "Z"
qui correspond en abscisses aux différents couples rédox qui interviennent et
en ordonnées aux valeurs des potentiels rédox de ces couples (voir annexe 5). Les flèches droites correspondent au trajet des
électrons.

D'après
"Principes de Biochimie" Horton et al. (1994)"
On
a regroupé dans le tableau ci-dessous quelques caractéristiques des principales
molécules ou des principaux complexes qui interviennent dans ce schéma en
"Z" :
|
-
Complexe d'oxydation de l'eau (COE) Ce
complexe est encore appelé complexe Z ou complexe tyrosine Z. C'est
en fait une paire d'acides aminés tyrosine/histidine résidu d'enzyme de ce
complexe qui est le donneur primaire d'électrons du PS II. |
C'est un complexe enzymatique
contenant Mn et Ca de formule empirique : Mn4CaOxCl1-2(HCO3)y
Il est étroitement associé à PS II. L'équation d'oxydation de l'eau
s'écrit :
Chaque électron cédé sert à
régénérer une molécule de chlorophylle. |
|
-
Les phéophytines Ce
sont des molécules de chlorophylle sans l'atome central de magnésium. On
distingue comme pour les chlorophylles 2 phéophytines
importantes : A
(ou α), B (ou β). C'est la A qui est la plus commune. Ces
molécules sont impliquées dans la photosynthèse au même titre que les
chlorophylles correspondantes. La
phéophytine a est le premier accepteur d'électrons
de la chaîne du PS II. |
|
|
-
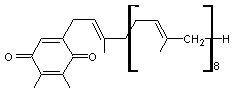
La plastoquinone Composé
quinonique de structure proche de celle des ubiquinones, que l'on
trouve dans les plastes des cellules végétales et qui participent à la
fonction chlorophyllienne. C'est Kofler
qui l'a décrite pour la première fois. Elle
se transforme réversiblement en plastohydroquinone
par réduction (ou inversement par oxydation). Elle
permet le transfert d'électrons de la quinone B au complexe b6f et un
transfert de protons du stroma au lumen. |
Plastoquinone
Plastoquinone Plastohydroquinone
|
|
C'est une métalloprotéine, petite
protéine de 10 kDa contenant du cuivre qui transfère les électrons du
complexe b6f au photosystème I. |
|
|
|
C'est
une oxydoréductase qui catalyse la réaction : Plastohydroquinone + 2 plastocyanines
oxydées + 2 H+ = Plastoquinone + 2 plastocyanines
réduites + 2 H+
|
|
-
La phylloquinone C'est
un accepteur d'électrons. |
|
|
-
Les centres [Fe-S] |
Ce sont des groupements
prosthétiques de très nombreuses protéines ; ils sont ici impliqués dans le
transfert des électrons.
|
|
-
La ferrédoxine |
Cest une protéine fer-soufre qui participe
aussi au transfert d'électrons.Elle contient des clusters Fe2S2
composés de deux cations de fer et deux anions sulfure. Elle intervient dans les réactions de
photophosphorylation c'est-à-dire la formation d'ATP à partir d'ADP à
laquelle participe une ATP synthase.
|

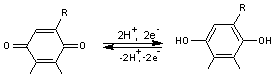
Remarque : Outre ce transfert acyclique
d'électrons qui voit la mise en jeu des deux systèmes PS I et PS II et qui
aboutit à la formation d'ATP et de NADPH, il existe un transfert cyclique
d'électrons qui n'intéresse que le système PS I et qui aboutit à la formation
d'ATP seulement.
3) La synthèse des
glucides ; Le cycle de Calvin
Le
cycle de Calvin correspond à une série de réactions biochimiques indépendantes
de la lumière (phase obscure), qui utilisent l'énergie chimique de l'ATP et du
NADPH (voir annexe 1) formés dans la phase claire
précédemment décrite, pour transformer le CO2 en glucose et autres
molécules organiques utilisées par l'organisme.
Ce
cycle comprend 3 étapes
|
|
|
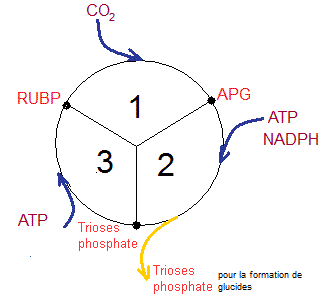
|
D'après
Etapes du cycle de Calvin Université Pierre et
Marie Curie (Paris – Jussieu) |
Etape
1
Incorporation
du CO2 qui, en réagissant avec le RUBP,
donne 2 molécules d'APG sous forme de glycérate.
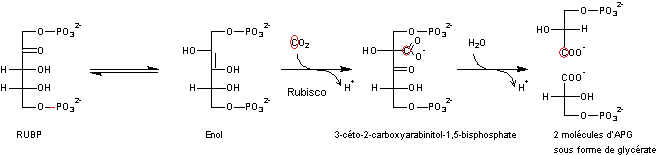
Remarque
: Le Rubisco pour Ribulose-1,5-bisphosphate carboxylase/oxygénase est un
enzyme, très abondant sur Terre, permettant la fixation du CO2 sur
le RUBP, première étape du cycle de Calvin de la photosynthèse. Il intervient
également dans la respiration (fonction oxygénase) des végétaux.
Etape
2
Réduction
de l'APG en trioses-phosphate
Elle
consiste en une transformation de l'APG en glucide par réduction de la fonction
acide en fonction aldéhyde
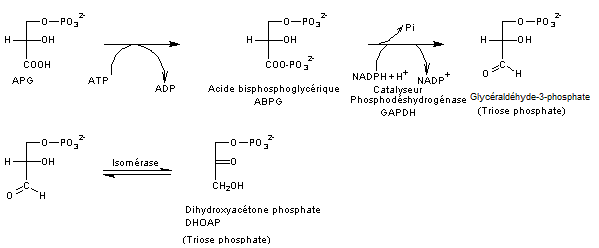
Etape
3
Il
y a régénération du RUBP à partir des trioses phosphate par les réactions
suivantes
- Réaction 1
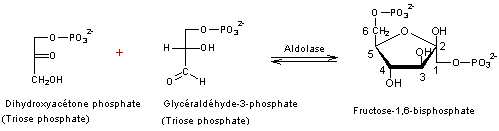
- Réaction 2
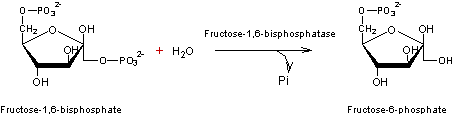
- Réaction 3
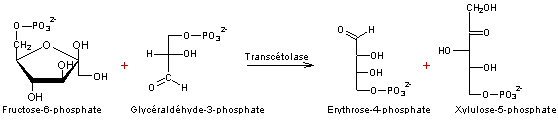
-
Réaction 4
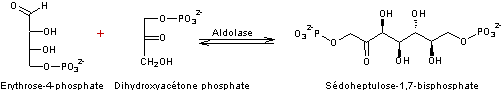
- Réaction 5
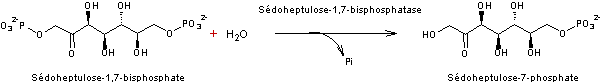
- Réaction 6
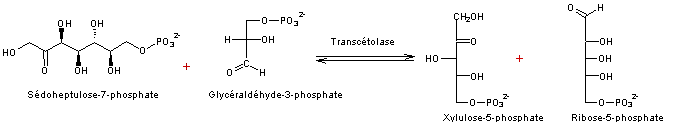
- Réaction 7

- Réaction 8
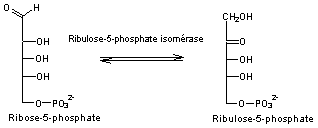
- Réaction 9
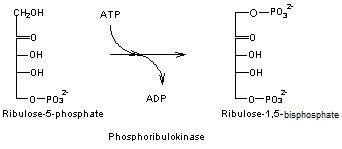
Le
Ribulose 1,5-bisphosphate correspond au RUBP.
Lors
de cette 3ème phase du cycle de Calvin on peut remarquer en
observant le devenir des glucides au cours des transformations, qu'à partir de
5 molécules de trioses (5xC3) on obtient 3 molécules de pentoses
(3xC5)
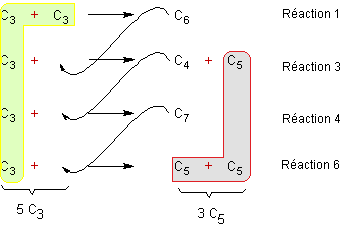
D'après
Ressources numériques en biologie rnbio.sorbonne-universite.fr
A
la lumière des équations ci-dessus, on peut réécrire le cycle de Calvin plus
complet et plus quantitatif :
|
|
|

E.Jaspard 2005
Globalement,
les 36 atomes de carbone obtenus à la fin de la première étape (30 provenant
des 6 molécules de RUBP et 6 de 6 molécules de CO2) se maintiennent
dans le cycle jusqu'à la 3ème étape ; la régénération du RUBP ne nécessitant
que 30 atomes de carbone, les 6 restants vont donner les glucides présents dans
l'équation bilan générale de la photosynthèse à partir des 6 molécules de CO2.
Il
existe trois types principaux de photosynthèse chez les plantes, qui sont la
photosynthèse C3, C4, et CAM.
Ces
différents types de photosynthèse correspondent à des mécanismes d'assimilation
du carbone légèrement différents, adaptés à diverses conditions
environnementales.
4-1) Photosynthèse
C3
La
photosynthèse C3 est le type le plus courant, se produisant chez la majorité
des plantes (environ 85 % des espèces). Elle tire son nom du fait que le
premier produit stable formé au cours de la fixation du carbone est une
molécule à trois carbones, le 3-phosphoglycérate (3-APG).
- Mécanisme: Lors de la photosynthèse C3, le dioxyde
de carbone (CO₂) est
fixé par l'enzyme RuBisCO (Ribulose-1,5-bisphosphate
carboxylase/oxygénase) pour former le 3-APG dans le cycle de Calvin. Ce
processus se déroule entièrement dans les cellules du mésophylle des feuilles.
- Conditions : La photosynthèse C3 est des plus efficaces dans des
conditions de faible intensité lumineuse, de températures modérées, et de
niveaux élevés de CO₂.
Cependant, elle est moins efficace en conditions de forte luminosité, de
sécheresse, ou de températures élevées, car la RuBisCO
a tendance à fixer l'oxygène plutôt que le CO₂, entraînant dans un processus
appelé photorespiration, une perte d'efficacité.
4-2) Photosynthèse
C4
La
photosynthèse C4 est une adaptation trouvée chez certaines plantes,
principalement des graminées comme le maïs et la canne à sucre, qui vivent dans
des environnements chauds et secs. Ce type de photosynthèse est plus efficace
que la C3 dans ces conditions.
- Mécanisme : Le mécanisme C4 commence dans les cellules du mésophylle
des feuilles. Le CO₂
atmosphérique est d'abord fixé par une enzyme appelée phosphoénolpyruvate
carboxylase (PEPC), qui a une plus grande affinité pour le CO₂ que Rubisco et n'interagit pas avec
l'oxygène.
1. Le CO₂ est fixé au phosphoénolpyruvate
(PEP), une molécule à trois carbones, pour former un composé à quatre carbones,
l'oxaloacétate.
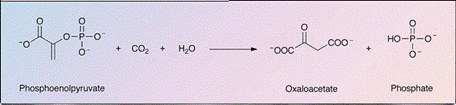
2. L'oxaloacétate est rapidement
converti en malate ou en aspartate, selon les espèces de plantes. Ce composé à quatre
carbones est ensuite transporté vers les cellules de la gaine périvasculaire,
où se déroule la seconde phase de la photosynthèse.
3. Une fois dans les cellules de la
gaine périvasculaire, le malate (ou aspartate) est décarboxylé pour libérer du
CO₂. Ce processus augmente localement
la concentration de CO₂,
ce qui réduit considérablement la probabilité de photorespiration
4.
Le CO₂
libéré est ensuite fixé par l'enzyme Rubisco dans le cycle de Calvin, où il est
utilisé pour produire des sucres.
5.
Après la décarboxylation, il reste une molécule à trois carbones, le
pyruvate. Ce pyruvate retourne dans les cellules du mésophylle, où il est
phosphorylé pour reformer le phosphoénolpyruvate (PEP), utilisant l'énergie
sous forme d'ATP pour permettre une nouvelle fixation du CO₂.
6.
Le cycle recommence avec la fixation du CO₂ au PEP par la PEPC, ce qui permet
aux plantes C4 de maintenir un flux continu de CO₂ vers le cycle de Calvin, même
lorsque les stomates sont partiellement fermés pour limiter la perte d'eau.
- Conditions : La photosynthèse C4 est avantageuse dans les
environnements chauds, ensoleillés, et souvent secs. Elle permet aux plantes
d'utiliser l'eau et le CO₂
plus efficacement que les plantes C3.
4-3)
Photosynthèse CAM (Crassulacean Acid Metabolism)
La
photosynthèse CAM est un mécanisme adapté aux environnements arides, présent
chez les plantes succulentes comme les cactus et certaines plantes épiphytes.
- Mécanisme : Dans la photosynthèse CAM, la fixation du CO₂ est séparée dans le temps. La nuit,
les stomates des plantes CAM s'ouvrent pour absorber le CO₂, qui est fixé par l'enzyme PEP
carboxylase en un acide organique, généralement le malate, qui est stocké dans
les vacuoles des cellules. Le jour, les stomates se ferment pour conserver
l'eau, et le malate est converti en CO₂,
utilisé ensuite dans le cycle de Calvin.
- Conditions : Ce type de photosynthèse est particulièrement avantageux
dans les environnements très arides, où la conservation de l'eau est
essentielle. Les plantes CAM peuvent ainsi continuer à fixer le CO₂ tout en minimisant la perte d'eau.
4-4) En résumé
-
Photosynthèse C3 : Fixation directe du CO₂
par la RuBisCO, efficace dans des conditions
tempérées.
- Photosynthèse C4 : Fixation du CO₂
par la PEP carboxylase et concentration du CO₂, minimisant la photorespiration,
efficace dans des environnements chauds et ensoleillés.
- Photosynthèse CAM : Fixation du CO₂
la nuit et séparation temporelle du cycle de Calvin, adapté aux environnements
arides pour économiser l'eau.
Ces adaptations permettent aux plantes de survivre et de prospérer dans une
grande variété d'environnements, en optimisant la photosynthèse selon les
conditions locales.
Reproduire
ce que la nature fait de façon simple, en apparence, est un chemin de recherche
sur lequel de nombreux scientifiques se sont engagés.
Il
s'agit d'utiliser de l'énergie lumineuse pour déclencher des processus
conduisant à l'oxydation de l'eau ce qui permet grâce aux électrons produits de
réduire CO2 et d'aboutir à des molécules organiques qui par exemple
serviront de carburant ou plus simplement à du dihydrogène.
De
nombreuses voies ont été et sont explorées, au début en utilisant des métaux
plus ou moins rares et précieux ; la tendance actuelle étant d'éviter ceux-ci.
On
donne ci-dessous deux exemples.
Les
progrès sont lents car la voie est difficile.
5-1)
Une piste
Les processus mis en jeu pour produire les glucides sont,
nous venons de le voir, nombreux et complexes ; dans l'une des étapes
importantes intervient un composé quinonique qui se
trouve dans les plastes des cellules végétales, la plastoquinone.
Elle se transforme réversiblement en plastohydroquinone
par réduction (ou inversement par oxydation), comme cela a été vu plus haut.
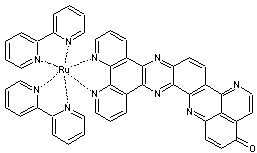
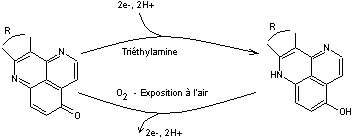
Artificiellement, par analogie, on peut retrouver cette
fonction grâce à un photosensibilisateur
qui permet une photoaccumulation de charges par un
complexe du ruthénium.
5-2)
Des travaux récents
Des travaux sur la photosynthèse artificielle primés par la
Royal Society of Chemistry
Une équipe internationale de scientifiques a reçu le prix John Jeyes de la Royal Society of Chemistry pour un système
produisant des carburants à partir d’eau, de dioxyde de carbone et de lumière
solaire.
|
|
|
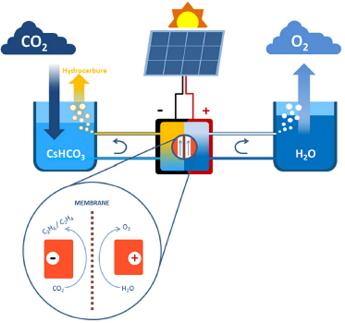
Schéma de principe
du système de photosynthèse
artificielle détaillé
dans “Low-cost high-efficiency system for solar-driven conversion of CO2 to
hydrocarbons” Tran Ngoc Huan et al,Proceedings
of the National Academy of Sciences May 2019, 116 (20)" |
Interview avec Martin Foldyna,
chargé de recherche au CNRS rattaché au Laboratoire de physique des interfaces
et couches minces.
"Avec vos collègues à travers le monde, vous avez
développé un système de photosynthèse artificielle. De quoi s’agit-il ?"
Martin Foldyna : La photosynthèse est un processus qui permet aux plantes
et à d’autres organismes vivants de convertir l’énergie solaire en énergie
chimique. Par exemple, les plantes peuvent réduire le dioxyde de carbone (CO2)
en sucre, qu’elles utilisent comme carburant. La photosynthèse artificielle
cherche à imiter ce processus afin de fabriquer de l’hydrogène et des produits
chimiques à haute valeur ajoutée, tels des hydrocarbures, à partir de CO2,
d’eau et d’énergie solaire. Ces « carburants solaires » pourraient ensuite être
utilisés et contribuer à une consommation énergétique plus soutenable. En
elle-même, la photosynthèse artificielle n’est pas nouvelle, mais dans notre
travail qui a été publié en 2019 dans la revue PNAS, nous avons réalisé un système
efficace à bas coût.
"Comment fonctionne ce système ?"
Martin Foldyna : Notre machine comprend deux parties : un module
photovoltaïque et un module électrochimique. En laboratoire, nous illuminons le
module photovoltaïque par un éclairage semblable à la lumière solaire. Cette
lumière est convertie en électricité, qui alimente en tension et en courant le
module électrochimique, lequel est plongé dans de l’eau enrichie en CO2. Ce
module est composé de deux électrodes en oxyde de cuivre, un matériau qui agit
comme catalyseur pour les réactions chimiques. Ces réactions décomposent l’eau
en oxygène et hydrogène. Mais elles produisent également des composants à base
de carbone, comme l’éthane et l’éthylène. Résumé ainsi, cela semble facile.
Cependant, la chimie en jeu n’est pas triviale. De plus, même si produire ainsi
de l’hydrogène est relativement facile, le rendement global que nous avons
obtenu pour les hydrocarbures atteint 2,3 %, ce qui est déjà un très bon
résultat.
"Comment avez-vous travaillé ?"
Martin Foldyna : Il s’agit d’un vrai travail collaboratif. Le but consistait
à faire un système à la fois efficace et peu onéreux, parce que ce dernier
point essentiel pour envisager des applications pratiques. Nos collègues
chimistes du Collège de France ont conçu un catalyseur original pour les deux
électrodes : un oxyde de cuivre dont la structure à l’échelle microscopique est
dendritique, c’est-à-dire ramifiée. Cela donne aux électrodes une grande
surface, ce qui bénéficie aux réactions. De plus, le cuivre est un matériau
plus abondant et moins cher que d’autres catalyseurs tel l’iridium. Les
cellules photovoltaïques ont été fabriquées à l’Ecole polytechnique de Turin,
en Italie. Elles sont constituées de pérovskite, une famille de matériaux dont
les propriétés promettent beaucoup pour les applications solaires à bas coût.
Au Laboratoire de physique des interfaces et couches minces (LPICM*), j’ai été
impliqué dans l’intégration du système dans son ensemble : le faire fonctionner
et mesurer ses performances sous un éclairage solaire calibré. En particulier,
il fallait assembler plusieurs cellules pour réaliser le module photovoltaïque,
ce qui était un défi car les perovskites sont
fragiles et leurs performances varient au cours du temps. Au bout du compte, je
pense que le fait d’avoir développé un système complet et d’avoir démontré ses
performances en lumière solaire a été apprécié par le jury de la Royal Society
of Chemistry.
"Le projet se poursuit-il ?"
Martin Foldyna : Oui, de mon côté, je continue de travailler avec
certaines personnes de cette collaboration. Nous voudrions réaliser une «
feuille artificielle », c’est-à-dire un système vraiment tout-en-un qui puisse
être directement plongé dans l’eau, illuminé par la lumière solaire et qui
collecte les carburants solaires produits sous forme gazeuse. Mais les perovskites résistent mal à l’eau. C’est pourquoi je
travaille sur une alternative : des nanofils à base
de silicium. La technologie qui sert à faire croître ces nanofils
est semblable à celle utilisée pour les films minces, ce qui permettrait de
réaliser des électrodes efficaces, avec une grande surface, et toujours à bas
coût. Nous possédons une grande expertise sur cette technologie au LPICM et
nous sommes en train de soumettre une publication montrant la production
d’hydrogène par une telle feuille artificielle, sans aucune connexion externe.
Interview extraite du site
https://www.polytechnique.edu/fr/content/des-travaux-sur-la-photosynthese-artificielle-primes-par-la-royal-society-chemistry du 7 juillet 2021.
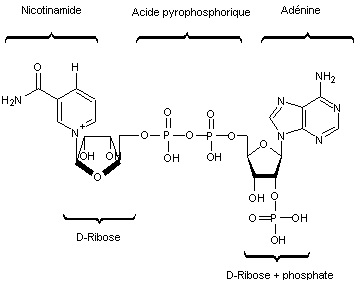
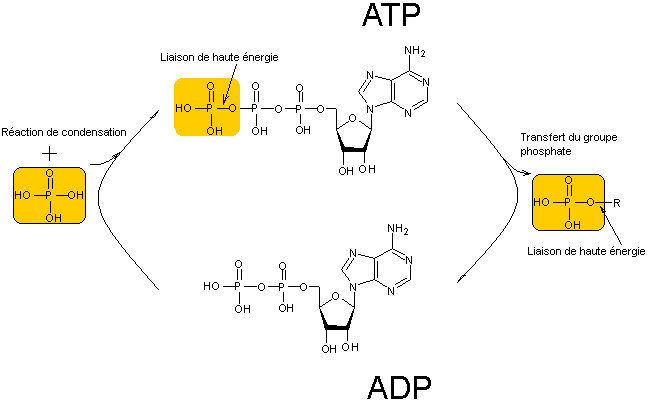
NADP+/NADPH
ATP/ADP
-
Coenzyme Nicotinamide – Adénine – Dinucléotide Phosphate (NADP+/NADPH)
|
NADP+ |
|
NADPH |
Il s'agit d'un couple
d'oxydo-réduction NADP+/NADPH dont la forme oxydée est NADP+ et
la forme réduite NADPH.
NADPH joue le rôle de réducteur dans
les réactions anaboliques avec la transformation

L'action de l'enzyme NADPH oxydase
couplé à NADPH permet par exemple la synthèse d'un dérivé réactif de l'oxygène
(DRO), l'anion superoxyde qui peut endommager les structures cellulaires.
-
ATP/ADP
|
ATP |
C'est l'acide adénosine triphosphorique, en pratique l'adénosine triphosphate
(ATP) qui joue un rôle fondamental pour les échanges d'énergie dans les
cellules.
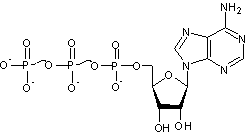
(Les liaisons ~ correspondent aux
liaisons riches en énergie dans la molécule).
|
ADP |
Adénosine Diphosphate
|
ATP/ADP |
Passage de l'ATP en ADP et
régénération de l'ATP
Chloroplastes, thylakoïdes, stroma
Le cytoplasme d'une cellule chlorophyllienne,
région située entre la membrane plasmique et le noyau (d'une cellule eucaryote)
est constitué de 80% d'eau et de différentes substances dont les chloroplastes
(plusieurs dizaines par cellule).
La taille des chloroplastes est de
l'ordre du micromètre ; un chloroplaste est délimité du reste de la cellule par
deux membranes constituant l'enveloppe et il contient des thylakoïdes (du grec thylakos,
sac et oides semblable), petits sacs empilés (l'empilement
s'appelle un granum) dont la membrane est riche en pigments absorbant la
lumière. Parmi ces pigments citons la chlorophylle, majoritaire, verte, mais
aussi des caroténoïdes jaunes ou orangés.
Entre ces thylakoïdes un fluide
aqueux, incolore, le
stroma où se produit la phase obscure de la photosynthèse.
On appelle lumen des thylakoïdes, l'intérieur de
ceux-ci.
Remarque :
Par souci de clarté d'autres structures présentes dans le stroma des
chloroplastes comme l'amidon, des structures lipidiques, de l'ADN, de l'ARN,
des ribosomes n'ont pas été représentés.
Caroténoïdes – Carotènes- xanthophylles
- Caroténoïdes
Ce sont des pigments constitués d'une longue chaîne
d'hydrocarbures polyènes ou de leurs dérivés oxygénés.
Le groupe chromophore
est fait de doubles liaisons C=C conjuguées. La majorité des caroténoïdes a
huit résidus isoprène, donc 40 atomes de carbones ; on peut aussi les rattacher
aux tétraterpénoïdes.
Parmi eux on peut distinguer les carotènes qui sont les
hydrocarbures, et les xanthophylles qui sont les dérivés oxygénés.
-
Carotènes
On peut les diviser en deux groupes, les alicycliques
α, β, γ carotènes et les aliphatiques par exemple le lycopène ou
le ζ-carotène.
-
Carotènes alicycliques
|
|
L' α-carotène Ce pigment jaune orangé constitue environ 15% des
hydrocarbures des carottes. |
|
|
Le β-carotène Pigment jaune-orangé. |
|
|
Le γ-carotène Pigment jaune orangé constituant environ 0,1% des
hydrocarbures des carottes Il ne possède qu'un cycle à une extrémité de la chaîne. |
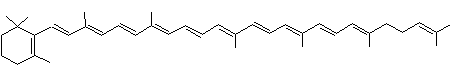
Toutes ces molécules ont comme formule C40H56
; ce sont des isomères.
Seuls les carotènes α et β apparaissent dans les
feuilles vertes.
-
Carotènes aliphatiques
|
|
Le Y,Y-carotène ou
trans-lycopène Comme les carotènes alicycliques il a comme formule C40H56
mais sa structure est linéaire. Il donne à la tomate sa couleur rouge. On le trouve aussi
dans l'abricot, l'orange... |
|
|
Le ζ-carotène Sa structure se rapproche de celle du lycopène mais avec 4
hydrogènes supplémentaires (C40H60). |
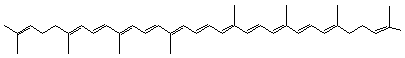

-
Xanthophylles
Ce sont des pigments jaunes ou jaunes-orangés constitués par
des dérivés oxygénés de carotènes.
Outre la lutéine citée dans le texte, on donnera l'exemple
de la Zéaxanthine.
|
|
La zéaxanthine On la trouve dans le maïs d'où son nom. Sa molécule est voisine de celle du β-carotène ou
deux groupes hydroxyles remplacent deux hydrogènes. C'est une molécule voisine et à action comparable à celle
de la lutéine dans l'organisme humain notamment au niveau de l'oeil. |
Chlorophylle A,
chlorophylle B, xanthophylles
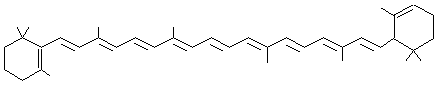
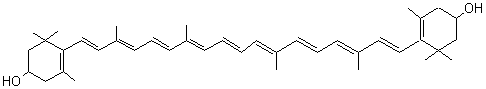
- Chlorophylles A et B
Les chlorophylles
constituent les pigments verts majoritaires.
Ce sont essentiellement les
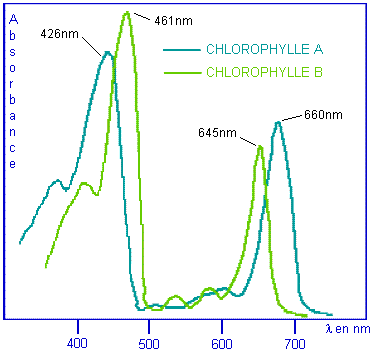
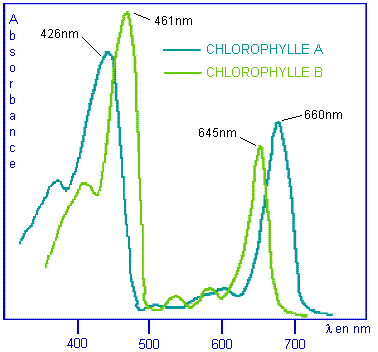
chlorophylles A et B qui absorbent la lumière à 426 nm (bleu) et 660 nm (rouge)
pour la chlorophylle A et à 461 nm et 645 nm pour la chlorophylle B.
Leur structure
correspond à un noyau tétrapyrrolique contenant un
atome de magnésium.
|
Structures
des chlorophylles A et B |
Courbes
d'absorption optique des chlorophylles |
|
Chlorophylle A : X = CH3 Chlorophylle B : X = CHO
|
|

- Xanthophylles
Les Xanthophylles (du
grec xanthon = jaune et phullon = feuille) sont des pigments jaunes ou orangés, dérivés oxygénés
des carotènes (voir annexe 3).
On peut citer la lutéine
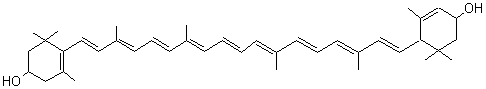
un pigment orangé présent
dans de nombreux légumes à feuilles vertes (brocolis, chou vert, épinards,
petits pois, endives crues....).
Les xanthophylles sont masqués par la chlorophylle dans les feuilles vertes.
valeurs des potentiels rédox
des couples intervenant dans la photosynthèse
|
Couples rédox |
E'0 (V) |
|
O2/H2O |
+0,82 |
|
P680/P680* |
+0,9 |
|
P680*/ P680 |
-0,8 |
|
Phéophytine (Red/Ox) |
-0,6 |
|
PQA-PQB
(Red/Ox) |
-0,2 |
|
PQres(Red/Ox) |
0 |
|
b6f(Red/Ox) |
-0,2 et +0,2 |
|
P700/P700* |
+0,4 |
|
P700*/P700 |
-1,3 |
|
A0 (Red/Ox) |
-1,0 |
|
Fd (Red/Ox) |
-0,42 |
|
NADP+/NADPH |
-0,32 |